
Home Page |
MICHIGAN FORESTS FOREVER TEACHERS GUIDE
| FOREST ECOLOGY BASICS |  |
A forest is a collection of biological organisms and non-biological factors. From an ecological perspective, the definition of a forest includes all these things, from the trees to the bacteria, and the soil type to the microclimates. See the "Tree Basics" page for more about the definition of a forest. Forest management systems are rooted in forest ecology (pun intended!).
There are three groups of concepts with forest ecology, or the ecology of just about any natural system. Each of the three groups of concepts interact with each other to variable degrees, at variable times, and in variable ways.
 |
|
Ideas on this page:
| Diversity Populations Communities Forest Layering Crown Cover Edge Effect Fragmentation Parcelization |
Snags Microenvironment Visual Quality Aesthetics Food Chains Nutrient Cycles Organic Matter |
Trophic Levels Weathering Hydrologic Cycle Temperature Humidity Succession Disturbance |
Composition has to do with species, taxonomy, and biological diversity. The number of species and how they relate to each other according to taxonomic classes is a reasonably straight-forward concept. Biological diversity, on the other hand, is a bit more slippery to wrap our minds around. At first, diversity sounds like a simple species count and relative abundance of each species. This is certainly a component of the diversity question. It's also the easiest to identify and study. However, the diversity of species is only one level of several.
| Species: the main category of taxonomic classification into which genera are subdivided, comprising a group of similar interbreeding individuals sharing a common morphology, physiology, and reproductive process. Note 1, there is generally a sterility barrier between species, or at least reduced fertility in interspecific hybrids. Note 2, the species is the basic unit of taxonomy on which the binomial system has been established; the lower taxonomic hierarchy is species, subspecies, variety, and forma. |
At the most fundamental level of diversity, there is genetic diversity. How many genes and pieces of genetic information are present in a forest? The chlorophyll gene, for example, is common throughout most of the plant kingdom (although there are several variants). Many other attributes or genetic characteristics are also quite common in a forest system. Vertebrates have far more genetic commonality than genetic difference. The loss of a species may not represent a loss of genetic diversity, only the loss of particular combination of genetic material. The raw material will probably remain in the biota. If you remove the word "the" from the English language, it would make our speech awkward, but it would not eliminate all words with the letters "t", "h", and "e".
Species diversity is the next level of diversity. These are the combinations of genes that we are most accustomed to dealing with in the life sciences and from legal perspectives (e.g. endangered species laws). Yet, the definition of the word "species" escapes a single, concise, universally-accepted agreement. Species abundance addresses the issue of how common a particular species is, often in the context of a particular region or ecosystem. There might be 100 species in a suite of characters. However, 90% of the biomass might consist of only 3 species. There may be a few species that are very uncommon, or have low abundance. It is usually the species with low abundance that we are concerned about from the perspective of potential species loss. It is usually the abundant species that we derive the most of economic base from. See the tree species diversity page for more information about Michigan forests.
A collection of individuals of a given species make up a population. The size, frequency, and distributions of populations are important elements of diversity. A Canada lynx might be listed as endangered in Michigan, but across its range it is a common animal. Sometimes populations on the edge of species range will display unique set of genes. Northern populations of animals tend to have larger sizes and shorter appendages. Flowering times of a tree species varies with climate conditions. The conservation of distinct populations may be important in some cases.
With a given ecosystem, populations interact with other. There are identifiable associations of species. These species associations are called communities. Community diversity is more difficult to measure in the landscape because there are usually a large number of components. To make identification possible, key species are used to describe a community, such as "northern hardwoods", which is defined by such tree species as sugar maple, beech, and basswood. However, northern hardwood associations in the western Upper Peninsula lose the beech component. Community descriptions have an inherent degree of variability across a large geographical region. This is another element of "diversity".
Lastly, there is something called "ecosystem diversity". A collection of communities and the association physical factors make up an ecosystem. Ecosystem diversity is commonly described in terms of biomes, eco-regions, and similar large-scale terms. However, ecosystems can also be quite small. The various communities within a rotting log are distinct from the surrounding forest. That rotting log, or all the rotting logs in a forest, can be considered an ecosystem.
While the above levels of diversity suggest a strong hierarchy, the classifications are less distinct in the natural world. A considerable amount of flexibility and "confusion" exists. Diversity is complex set of concepts, despite our understandable tendency to reduce it to a species level.
|
|
Activity Suggestion PLT Charting Diversity PLT The Fallen Log |
The structure, or "architectural" arrangement of a stand and forested region is important. Structure impacts wildlife habitat in a major way. It also influences light, water, and nutrient levels. These things, in turn, impact the trees and other vegetation. Structural components of a forest, or lack of a particular component, are not inherently "good" or "bad", or "natural" or "unnatural". Structure is an ecological feature of a forest that can be measured, and subsequently evaluated against a set of criteria. Seven elements of structure are discussed.
Vertical & Horizontal Arrangement
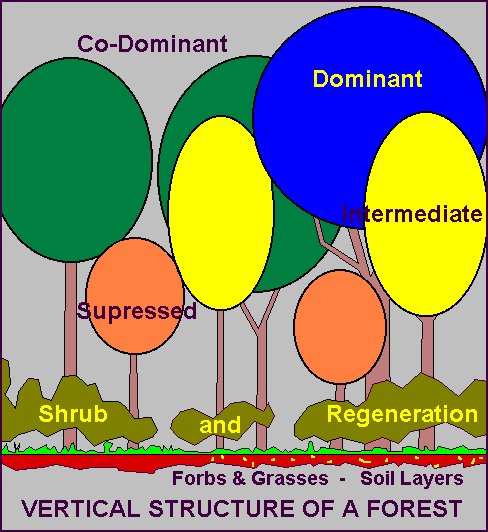 |
This is the physical arrangement of a forest; the different tree heights, "layers" of forest, and the continuity of branches from tree to tree. A forest with more structure generally has more habitat characteristics. A continuous forest offers transport routes for arboreal animals (animals that live in trees). "Crown cover" is the percent of the ground that has tree crown growing over it. A forest will have variable percentages of "holes" in the canopy. These "holes", or the amount of crown cover or crown closure, have important ecological ramifications.
Classical tropical humid rainforests probably have the most structure of any forest on Earth. Our northern temperate forests are different. Not all forest types have a full complement of layering (understory, shrub, mid-size trees, main canopy trees, really tall trees). Jack pine stands, for instance, generally lack much vertical structure, especially with the kinds of soils they typically grow on. Northern hardwoods, if managed accordingly, will have a well-developed structure. Left unmanaged, they tend to lose structure close to the ground.
Heterogeneity and Forest Density
The level of "heterogeneity" refers to how similar or different the parts of a forest are to each other. Diversity is a big part of this, but so is structure and other forest descriptors. Heterogeneity might be evaluated within a single stand of trees, or be assessed across a large landscape, such as a national forest or the eastern Upper Peninsula.
"Density" has to do with how many trees are in an area and how large the trees are. A thousand trees per acre may, or may not, be a lot of trees depending upon their size. A thousand seedlings are generally more than recommended, but the density is still low. 250 large, sawtimber-sized trees per acre would likely be a high density forest. Density is typically measured in units of square feet per unit area (acre) and is called "basal area" (link to the Forest Descriptor page in the Tree Basics section).
 The design of a timber harvest can have significant on habitat quality. Within a given area, more or less "edge" can be created according to the management objectives of the forest stand. |
"Edge" refers to the transition zone between two different vegetation types. Some "edge zones" are sharp or narrow, such as the transition between a lake and the shoreline vegetation. Other transitions are more gradual, such as the change in forest composition up a slope in the Keweenaw Peninsula. These edge zones usually have representative species from both vegetation types, so they tend to be more diverse than either of the constituent types. Edge zones may also have species not found in either constituent type. In terms of broad-brush diversity, forest edge is a good thing, especially if higher numbers of species were the only measure.
The downside of forest edge is the affect on species that prefer or require "deep woods" conditions. The introduction of an open edge to a "deep woods" type forest will stress those species that don't benefit from edge effects. The ovenbird is often cited as an example of an animal that can only be found in undisturbed, mature forests. This conclusion is arguable, as ovenbirds can often be heard in other forest types. What the ovenbird really needs is protection from ground predators, as they build their nests on the ground. Deep, dark forests tend to have fewer species and do not support high numbers of large predators.
Islands and Fragmentation
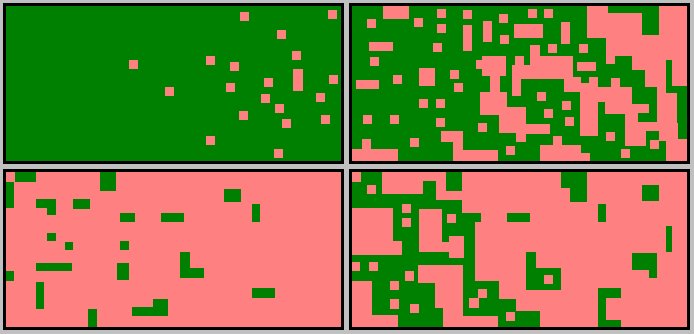 Forest fragmentation is the land use change from forest cover to a non-forest cover. This is represented in the above image starting from the upper left and moving clockwise. |
The process of changing a large forested area into an area of forest patches is called "fragmentation". These forest patches are referred to as "islands." The fragmentation of forest has important ecological impacts. These impacts are not necessarily "good" or "bad" but they are definable, at least in part.
Forest fragmentation should not be confused with forest "parcelization". Parcelization is an ownership phenomenon that often, but not always, translates into forest fragmentation. Parcelization has direct economic impacts, as well as potentially direct ecological impacts.
Mathematical theories relating biological diversity and fragmentation were promulgated by a man named E.O. Wilson. These theories were developed to explain ecological trends and patterns found among the islands of the South Pacific. The theory is called "island biogeography". Both the original theory and its subsequent application to continental situations remain highly controversial.
Dead Trees and Snags
Dead trees, both on the ground and standing, provide habitat elements for many species, particularly cavity nesters. Dead wood also provides habitat for a number of insects which, in turn, are important parts of some food chains. A standing dead tree is called a "snag". Large snags and fallen trees have more value than small ones.
"Snag management" means producing more snags in a forest where snags are uncommon. From a strict timber perspective, snags are trees that could have been merchantable had timber been the only management goal and management implementation was perfect. As a legacy of our forest history, many of our forests remain in younger, more vigorous age classes (although this trend is reversing). Snags become increasingly common as forests grow older. Forests of short-lived species already display an abundance of snags. The presence of snags in longer-lived forests can sometimes be accelerated through management. Snags in short-lived forests are usually retained during timber harvest.
Small areas within a forest environment that have markedly different characteristics are called "micro-environments". Examples might be rock outcrops, large rotting logs, pit-mound topography (tree tip-ups), springs and seeps, vernal (spring-time) ponds, or other features. These micro-environments sometimes harbor special sets of species, and occasionally endangered or threatened species. Seeds of some tree species may depend on micro-climates / micro-environments to enhance their reproduction success. Manipulation of the forest canopy through management practices alters micro-environmental conditions such as light, temperature, and humidity.
Appearance
The "appearance" of a forest is not really an ecological factor, but it has great influence on how forests are managed or not managed. Forest management or lack of management can have significant ecological impact. The appearance of a forest strongly influences public opinion and public policy of the "goodness" or "badness" of a particular forest practice. An entire of compendium of "pseudo-science" has been developed to support what is essentially an argument against forest practices resulting in poor appearance. Generally, balanced ecological and biological information is not considered. This phenomenon is not unique to natural resources, of course.
"Visual quality" is the term often used as an objective in forest management. "Aesthetics" is a misnomer that is also quite common. An aesthetic involves the appreciation of something. Appreciation has deeper meaning than mere visual appearance. A well-done and properly applied clearcut can have high aesthetic value, but rather low visual quality.
The "function" part of an ecosystem involves "how" things happen. It's equivalent to "economics" in our human society, or the themes of geography that involve the movement of goods and interactions between humans and the environment. How ecosystem functions are played-out can be highly complex, but the functions themselves are fairly easy concepts to understand. These areas of forest ecology are probably the least understood, but the most resilient.
Nearly all life on Earth is solar-driven. Plants capture solar energy and store it as chemical energy (photosynthesis). Animals eat plants to obtain this stored energy, among other things. Some animals eat other animals, for the same reasons. These threads of energy transfer are called "food chains". Energy can be likened to the currency that measures an ecosystem economy. Life bucks the laws of entropy. In this sense . . . life is not "natural", but eventually all the energy is dispersed.
 |
The "rule of 10 percent" says that only 10 percent of the energy in each transfer is actually captured. So, plants only capture about 10 percent of the solar energy available to them (actually, it's less than this). Herbivores capture only 10 percent of the energy stored in plants. And, the same is true "on down the line" of the food chain. Energy transfers occur between "trophic levels".
In addition to energy, a host of minerals and nutrients cycle through the biota. Understanding these cycles involves chemistry, biology, and physical geography. The most common elements are carbon, hydrogen, oxygen, phosphorus, potassium, nitrogen, sulfur, calcium, iron, and magnesium. A jingle to help remember these nine elements is "CHOPKNS CaFe Mg" or "see Hopkins Cafe, might good". There are another couple dozen or so elements needed, too.
How minerals and nutrients cycle through the biota varies considerably. Decomposers, soil type, water, and climate are determining factors. Cycles tend to be "open" in temperate zones, meaning nutrients are commonly lost to a system or a food web. A significant portion of available nutrients are found in "organic matter", or the layer of dead material on top of the soil surface. Tropical humid systems tend to be "closed" with very few nutrients slipping out of the system. Soils in these regions have very little organic matter.
The availability of a nutrient differs from the amount of a nutrient in the environment. For example, just because there is a lot of nitrogen in the atmosphere and the soil, doesn't mean it is in the form a plant can absorb. Nutrient availability varies with soil type, pH, and other factors. For more information about mineral & nutrient cycling, try the "cycles" page.
New minerals and nutrients are added to an ecosystem as rocks and soils are chemically broken down by weather and biological factors. "Parent material" is the rock or mineral source(s) from which soils are derived. The productivity of a particular soil is highly dependent upon the parent material in the area. Parent material rich in key elements will produce soils that support higher levels of biomass. Weathering is the prime source of "new" minerals and nutrients in an ecosystem.
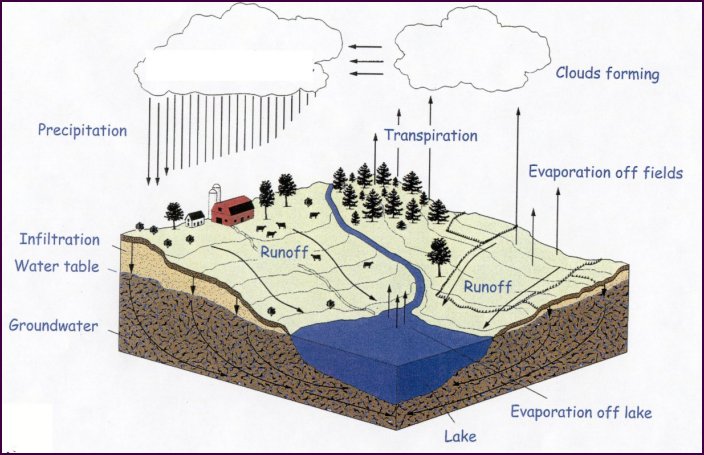 Image courtesy of the Michigan State University |
The "hydrologic cycle" is commonly taught throughout Michigan at the upper elementary and middle school level. The amount of water on the Earth is a fairly finite quantity. Where it occurs and how it cycles has a tremendous impact on the biota in an area.
Essentially, water cycles through the atmosphere, living matter, and the soil. Movement can be "stalled" by a number of features, such as lakes, underground aquifers, glaciers, etc. As water moves downward through soils, it usually takes soluble nutrients with it. Water movement is the main reason for the "loss" of nutrients in an ecosystem.
Like nutrients, water availability to plants also varies. Coarse soils, those with relatively large particle size and pore size, tend to hold less water. They are drought-prone and most tree species don't grow well on these soil types. Finer soils, such as silts and clays, have very fine particles and hold more water. However, during dry periods, they may be just as droughty as coarse (or sandy) soils because the drier these soils become, the tighter that water molecules "cling" to the soil particles. This cohesive property may be stronger than a root system's ability to pull the water molecules out of the soil.
For more information about the hydrologic cycle, try the "cycles" page.
Temperature & Humidity
Temperature and humidity play important roles in the transfer of materials throughout an ecosystem. They also have strong influences on "who grows where" and affect metabolic processes of both plants and animals.
Photosynthetic rates correlate to temperature. Higher temperatures increase rates, to a certain point, after which a plant can "burn out". Very arid environments cause plants to close their pores in order to conserve water. Less water inside the plants can slow photosynthesis to a crawl. Desert plants have special adaptations for dry conditions. So do many plants in our northern bogs and sandy outwash plains. And the temperature-humidity condition immediately above a forest canopy on a hot summer day can be every bit as hostile as that in a desert.
Successful germination of tree seeds and early seedling survival are quite sensitive to temperature and humidity conditions on the forest floor. For example, sugar maple seeds germinate in the spring, soon after snow melt, when the temperature is 34 degrees (F). If an early spring heat wave hits, germination for that year will be poor. On the other hand, yellow birch, a common associate of sugar maple, germinates best around 74 degrees. That's part of the reason why an unmanaged northern hardwood stand will often migrate towards a sugar maple monotype and a managed northern hardwood stand will encourage the regeneration of other northern hardwood tree species.
Succession and Disturbance
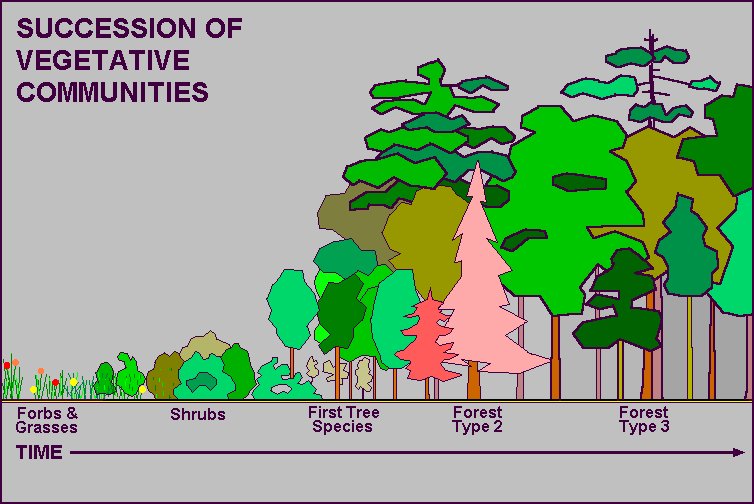 |
As the living (biotic) and non-living (abiotic) components of an ecosystem react with each other over time, the composition, structure, and function of a forest will change. This process of change is called "succession" which is more fully explained on the succession webpage. The classic concept of succession is the orderly and reasonably predictable path of vegetational change over time, from bare soil to mature forest. The early vegetation types (seres) are labeling as "pioneer" types. The final sere, if there is one, is called the "climax" type.
The progress of succession can be interrupted and set back by disturbances, usually the result of fire, wind, flood, or insect/disease epidemic, and nowadays by forest management. Disturbance might be on a grandiose scale, such a large wildfire, or a small scale, such as single large tree blown over in windstorm or killed by a lightning strike. Depending upon the severity of the disturbance, an earlier vegetational stage (or sere) will result, and succession will restart from that point.
Forest management is closely integrated with an understanding of forest successional pathways. Succession can be accelerated or slowed, depending upon the desired objectives for a particular stand and the natural capabilities of the site. Successional pathways vary according to many factors.
|
|
Activity Suggestion PLT Nothing Succeeds Like Succession |
 |
This website was developed and created by Michigan State University Extension for the teachers of the State of Michigan. |